|
訳者コメント: ゲノムスキャンのデータを比較的コンパクトにまとめてあります.Shaoらのゲノムスクリーンは本ホームページではまだ紹介しておりませんが,このレビューには印刷中として紹介してあります.入手できしだい論文コーナーに掲載します.グルタミン酸レセプターに関連する未発表データ(Jamain Sら)やDuke大学のR. Carneyらの未発表データも紹介してあります.このCarneyらの結果は,69例の女児自閉症例(症候的にはRett症候群ではない)の中に2人のMECP2変異例が見つかったとしてあり,自閉症におけるMECP2遺伝子の検討の最初の論文になりそうです.これも論文になりしだい掲載します.展望のところでは,自閉症の遺伝素因(相互作用を持つ複数遺伝子性)が特別なものではなく,一般ポピュレーションの中にも共通して存在するとする考え方が明記してあります.つまり,自閉症者がいる家系と,自閉症者がひとりもいない家系の間に遺伝的な本質的違いが実はなく,複数の関連遺伝子の単なる組み合わせで自閉症の遺伝的背景が形成されると考えることも可能なわけです.自閉症がヒトの性格と同じ遺伝的背景(QTLs)を持っているとする私どもの考えと一致する見解で,こういった考えが既に主流になってきていることを実感するレビューです(訳文中の図はコピーライトに配慮し全部書き直してあります). |
(概訳)
(概要)自閉症は1943年にdisorderとして始めて認識されてから,その原因については憶測の域を出ず,生物学的なものから心理学的なものまでいろいろな説が議論されている.1970年代から1980年代に行われた双生児研究が遺伝素因の関与を示す決定的な証拠を示した後,自閉症における病因研究は関与する遺伝素因を解明することに着目し始めた.自閉症者における染色体異常の同定やメンデルの法則に従った遺伝性症候群の発見は,ゲノムスクリーニング研究や候補遺伝子研究のデータとあいまって,自閉症スペクトル状態の背景となっている複雑な遺伝素因の存在をさらに示唆している.
(イントロ)自閉症は最初Leo Kannerが記載した.Kannerは小児精神科の創始者の一人で,アメリカにおける小児精神科の最初の教授であった(the Johns Hopkins School of Medicine).1943年,Kannerは11人の小児例を記載し,そのほとんどは男児で,社会的孤立を基盤としており精神遅滞と区別できる状態とされた.彼はこの症候群を「幼児自閉症:infantile autism」と命名した.その理由は症例にみられた社会的相互作用の欠如がEugene Bleulerが分裂病(統合失調症)における社会的興味の消失に使った基準に類似していたからである.1年後,ドイツのHans Aspergerは,同様のケースを記載し,その状態を「autistic psychopathy」と呼んだ.男女比は4:1で一貫して男性に多く,Kannerの基準を使った有病率は1万人あたり2-5人である.しかし,最近の診断基準の変化と診断基準の運用状況の変化の結果,有病率は増加している(ボックス1).
ボックス1 自閉症表現型
| 社会的発達の異常
生後一年間で,その後自閉症と診断される幼児はしばしばやり取り相互作用(to-and-fro interactions)への興味や擁護者への執着の発達に障害がある.その後,彼らは親に対する執着は発達するが,他の児への興味はほとんど育たない.青年期と成人期にはしばしば社会的な相互作用に興味を持つが,社会的活動の開始はぎこちない.アイコンタクトが乏しく,ぎこちないあいさつで,社会的なちょっとした会話が欠如しており,他の人にとっては全くおもしろくないトピックスを力説する. 言語発達の異常
融通の利かない反復性の行動
徴候および症候中の発達変化
神経病理および病態生理
有病率 |
カナーもアスペルガーも両者共,オンセットが早期であることを根拠に,自閉症が神経病理学的オリジンを持つことを論じた.しかし,1950年代には,この考えは疑問視された.なぜなら,自閉症児はコミュニケーションができないけれども,ほとんどの神経学的異常のある児に典型的な身体形態異常(dysmorphic features)もないようであり重篤な出生時外傷の既往もなく,「知的であるようにみえる(seemed intelligent)」ためである.また,両親はしばしば社会的には控えめであることもこの考えが疑問視された理由である.この時点でアメリカ精神科学会においては全ての精神科的障害が親の子育てにおける問題や早期人生の経験に起因するものと考える傾向が一般的になっていた.ゆえに,親にみられた社会的寡黙が自閉症の明らかな原因であるとする見解が出された.社会的寡黙性は父親においてより一般的であるにもかかわらず,母親は「冷蔵庫(のように冷たい)母」とレッテルをはられた.結局は,Bernard RimlandとMichael Rutterによって仮説が提出された後,自閉症児の両親は非自閉症児のコントロール児の親と比較して子供の育て方に何の違いもないことが経験的に立証され,そしてさらに自閉症の神経生物学的解釈が説得力を持つようになった.
このレビューでは,自閉症が遺伝的な状態である証拠を示し,可能性のある遺伝学的メカニズムの理論を紹介する.それから,自閉症遺伝子を持つと思われる染色体を同定することを目的とした遺伝学的研究について議論する.これらの遺伝学的研究には,ゲノムスクリーン研究,染色体検査所見やメンデルの法則に従う疾患との関連とその病態生理などから集められた手がかりが含まれる.最後に自閉症に関連する遺伝子の同定を促進する可能性のあるいくつかのアプローチについて述べる.
遺伝学的原因の証拠
カナーが最初に記載してから,自閉症の原因に遺伝が重要であることが認められるまでに20年以上も要した.最初の頃に報告されたケースでは,多くて2-3%の家系が複数の自閉症児を有していた.Rutterは,一般頻度を1万人あたり2-5人として,(家系内)重複例の頻度が偶然の50-100倍であると記載した.最初の双生児研究の報告が,二卵性一致率に比べ一卵性一致率が有意に高いことを明らかにした時,研究者達は単純なメンデルの遺伝パターンに合わないけれども,自閉症の遺伝学的基盤が存在することを示唆した.
双生児研究:自閉症に関する3つの疫学的双生児研究において,片方または両方とも自閉症の双生児で特定の地域に住んでいるケース全員が調査された.宣伝でボランティアをつのると,一卵性双生児例が集まりやすくまた一致例(2人とも自閉症)が集まりやすいバイアスがあるが,この方法はサンプリングパイアスを減らすことができる.
自閉症双生児研究はまた,同性の双生児ペアのみをサンプルとしている.これは自閉症においては男児例が多いため,男女ペアの二卵性双生児例では不一致率が上がるからである.しかし,3つの研究の対象サンプルを全部たしても66組にすぎず,36組の一卵性ペアと30組の二卵性ペアである.二卵性一致率が0%であるのに対し,一卵性一致率の平均は70%であった.この二卵性一致率はサンプルサイズが小さいことによる結果であろう.
一卵性双生児ペアにおける自閉症の一致率は出生前または周産期の障害を共有することでは説明することができず,従って一卵性一致率対二卵性一致率の比が大きいことは自閉症の遺伝性が高いことを示している.家系内再発生率(recurrence risk)と一卵性:二卵性一致率比から計算される遺伝性推定値は90%以上である.しかし,このことはそれぞれの表現型の違いに環境の影響がないことを必ずしも意味していない.単に,研究されたポピュレーションにおいて,環境によるバリエーションがほとんど存在していないことを意味しているのかもしれない.
自閉症またはより軽度の認知障害や社会性障害からなる表現型に関して一致率を検討すると,一卵性一致率は82%,二卵性一致率は10%までとなる.自閉症不一致例の一卵性双生児ケースにおいて,自閉症ではない方の多くは言語性の学習障害を有しており,また何人かは社会的に寡黙である.この自閉症不一致例の非自閉症者の社会的寡黙は,その双生児ケースが大人になってから再調査された場合によりはっきりしている.
自閉症者の非自閉症家族の研究:双生児不一致例の非自閉症者が類似した認知障害や社会性障害を持っているとしたFolsteinとRutterの発見は予想されるべきであったかもしれないが,予想されていなかった.1957年になってやっと,Leon Eisenbergは自閉症児の父親の多くが融通の利かなさと社会的相互関係への興味の欠如などの普通でないパーソナリティー形質を持っていると報告した.これらの父親達は結婚しているわけで,適切に社会生活が送れることが明らかな多くは成功した専門家であったが,自分の時間を過ごす時には一人で活動することを好み,ルーチンにこだわる傾向があった.このEisenbergの論文は実際,自閉症の原因が「冷蔵庫のような母親」であるとした精神分析医による予想が流布したことに対して出されたわけである.Eisenbergはもしどちらかの親がこのような状態であるとしたら,それは父親であろうと指摘した.
FolsteinとRutterの双生児研究の後,いくつかのグループが家族研究を再開したが,今度は遺伝素因を想定しており,その情報を使い自閉症が遺伝する遺伝学的メカニズムを確立するためのゴールを設定していた.全体としては,親と兄弟の研究では,いくつかの特徴がコントロールの親に比べ自閉症児の親でよりしばしば見られることがはっきりと示された.これらは,社会的寡黙傾向,コミュニケーション障害(言語の実用的側面の障害),ルーチンへのこだわりと変化にたいする抵抗などを含んでいる.これらの3つの形質は概念的には自閉症の診断基準と同じであるが,非常にマイルドでありより広い自閉症表現型(broader autism phenotype)として知られている.このような親は通常は機能的障害を伴わず(機能的障害を伴う可能性はあるが),到達度が高いことにむしろ関連している場合がある.FolsteinとRutterは,このような側面が遺伝的自閉症易罹患性の結果であることを提案した.
会話のオンセットが遅かったり,読みの障害があったりすることも,繰り返すうつ病や不安障害,血小板セロトニンの増加,頭周囲長の増加などと同じように,自閉症者の家族メンバーによく見られる.精神発達遅滞は自閉症児の兄弟において多いということはないが,例外的には自閉症の一部としてはあり得る.
部分的には家族研究を基盤として,そして自閉症と何人かの児にみられるより軽度な社会性障害の間の著明な類似性にも基づき,自閉症は現在,類似性を持ちおそらく遺伝的に関連している障害のスペクトルを包括するものとして考えられている.このスペクトルはけいれん発作を伴い会話がまったくない著明な精神発達遅滞者や,より典型的な自閉症,さらに正常な知性を有し構造的な言語能力のあるよりマイルドなタイプのアスペルガー症候群,そして自閉症発端者の家族メンバーの中に見られるさらによりマイルドなより広い自閉症表現型を包括する.
自閉症の遺伝学的メカニズム
罹患者に自閉症が通常より高率にみられるいくつかの異なる遺伝的疾患が知られている.また,染色体異常に関連する自閉症のケースもたくさん報告されている.これらのケースは後述するが,時にいろいろな発見を導く意味で重要であり,最近の疫学的研究の結果である自閉症の有病率の増加の一つの重要な原因である.
自閉症の背景となる遺伝学的メカニズムを発見するための研究は,特発性とされるケースのみを対象としてきた.原因不明の特発性にどういうケースが含まれるかは時代と共に変化してきたことを理解することは重要である.Kannerにとって,特発性例は原因不明というわけではなく,原因が不明であっても神経学的症候群を意味するかもしれない形態異常症候がない場合である.自閉症ケースの最近の解析は,遺伝学的研究におけるこういうアプローチ(特発性に限定して研究すること)に価値があることを示している.マイナーな先天的奇形がないケースに限定しても,男女比はほぼ10:1であり,家系内における自閉症重複発生率もかなり高い.カナーのケース群でも同じことが言える.最近の研究では既知の原因があるケースを除外しているが,軽度で原因不明の形態異常症候を伴ったケースは除外していない.
検査者によって特発性と判断される自閉症の場合,最もひかえめな遺伝モデルはいくつかの遺伝子がお互いに相互作用を持って自閉症表現型が誘導されるとするモデルである.家族歴データの解析では,Andrew Picklesらは単一遺伝子座性を否定し,2個から10個の遺伝子座が関与する複数遺伝子座モデルである非単一モデルを想定し,相互作用を有する(エピスターシス)3つの遺伝子座の関与が最も考えられるとした.異なるアプローチを使い,Paul Van Eerdeweghは同じような結論を得た.彼は,相互作用を持つ3個から6個の遺伝子座の関与がデータと矛盾しない最もシンプルなモデルであると結論した.自閉症表現型にそれぞれが寄与する3個または4個の遺伝子を親から受け継いだ児において自閉症が起こると考えることができるかもしれない.このモデルでは自閉症の原因となる遺伝子がいつも同じ3つまたは4つの遺伝子であるのか,あるいは原因遺伝子群の中から3つまたは4つの遺伝子が組み合わさって存在した時に自閉症になるのかどうかに結論がでていない.兄弟例における兄弟間の重症度に多様性があることや,自閉症発端者から集められた家族サンプルにおいて家族メンバーの中にアスペルガー症候群や他の軽症自閉症様表現型が含まれている事実は,このようなモデルで説明できる.自閉症発端者の親族におけるよりマイルドな表現型は,自閉症の背景となる遺伝子の一部(サブセット)の遺伝を反映しているのもしれない.
このモデルはもっともひかえめなモデルであるが,単なる一つのモデルに過ぎず,他の可能性も存在する.現在特発性と判断されている全てのケースに関連する単一のモデルはないことが一番考えられる.家族内における重症度のスペクトルを説明するもう一つの可能性は,言語障害または社会的寡黙性の遺伝的背景が合併し,環境リスクファクターまたは免疫原性リスクファクターによる追加影響(セカンドヒット)があって自閉症が起こるという可能性である.
免疫原性:かなりの間接的証拠が自閉症における自己免疫的機序の可能性を示唆している.コントロール発端者の家系に比べて,自閉症発端者の家系においては,自己免疫疾患がより多いことを一つの論文が報告している.2-3の論文は,自閉症児または彼らの母親の中にはMHC遺伝子座におけるハプロタイプが自己免疫病態に関連している可能性のある型であった.2つの報告において,コントロールよりも自閉症児の方がより多く,myelin basic proteinやneurofilament proteins,そしてvascular endotheliumを含む脳の組織やタンパク質に対する自己抗体が検出された.免疫原性を示す所見は研究間で結果が異なっており,ほとんどの報告が単一のラボからの少数サンプルでの結果を記載している.これらの所見の多くは,おそらくMHC分子の機能的欠質で説明しえるのであろう.しかし,下述するゲノムスクリーンの結果では,第6染色体短腕上のMHC遺伝子座の近傍には小さなシグナルさえ検出されていない.にもかかわらず,自閉症の原因における自己免疫の役割を支持する傍証が十分に存在しているため,他の遺伝子上の免疫関連遺伝子の検討を含む,より大規模で十分なコントロールを設定した研究がさらに行われることが必要である.
ゲノムスクリーン研究
いくつかのゲノムスクリーン研究が現在既に発表されており,かなり多数の示唆的連鎖シグナルが報告されており(表1,図1),論文間で共通する結果はわずかに2-3の部位である.それぞれの研究グループは異なる解析方法を使っており,そのために連鎖の証拠の絶対価値は比較することが困難である.また,いくつかの遺伝子が関与する(oligogenic)疾患においてシグナルの局在を決定することは極端に困難であり,そのため一つの所見に再現性があるかどうかもしばしばはっきりしない.このパターンは,家族研究データから示されたモデル(エピスターシスを伴ったいくつかの遺伝子の関与)に矛盾しない.もっとも,ゲノムスクリーン研究ではそのモデルを検証できたわけでもなく,単一モデルの存在さえも確認できていないのであるが.4つの研究グループはX染色体上に小さなシグナルを検出してはいるものの,ゲノムスクリーン研究によるほとんどの連鎖シグナルは,常染色体上にある.自閉症の男女比が4:1であるとして(特発性ケースでは男性例がさらに多いが),自閉症の原因におけるX染色体の役割はこれまでに検討されてきた.しかし,広げて家系を検討しても,自閉症やより広い表現型の父から息子への伝播と同じ様に,X染色体との連鎖関係は遺伝的多型のわずかな部分だけを説明するであろうことが示されている.
表1 ゲノムスクリーン研究
| 研究グループ | 年 | 兄弟ペア数 | 染色体領域 | マーカー | 最高LODスコア |
| IMGSAC | 1998 | 87 | 7q
16p 4p |
D7S530/D7S684
D16S407/D16S3114 D4S412 |
MLS 2.53
MLS 1.51 MLS 1.55 |
| CLSA | 1999 | 75 | 13q
13q 7q |
D13S217/D13S1229
D13S800 D7S1813 |
MMLS/het 2.3
MMLS/het 3.0 MMLS 2.2 |
| Philippe | 1999 | 51 | 6q | D6S283 | MMLS 2.23 |
| Risch | 1999 | 147 | 1p | D1S1675 | MMLS 2.15 |
| Buxbaum | 2001 | 総数95
狭義49
|
2q
2q 2q 2q |
D2S364/D2S335
D2S364/D2S335 D2S364/D2S335 D2S364/D2S335 |
HLOD 1.96
NPL 2.39 HLOD 2.99 NPL 3.32 |
| Liu | 2001 | 総数118
狭義75
|
5q
Xqter 19p Xqter 16p 19q 5q |
D5S2494
DXS1047 D19S714 DXS1047 D16S2619 D19S587/D19S601 D5S2488 |
MMLS 2.55
X-MLS 2.56 MMLS 2.53 X-MLS 2.67 MMLS 1.93 MMLS 1.70 MMLS 1.63 |
| IMGSAC | 2001 | 総数152
狭義127 |
2q
7q 16p 17q 2q |
D2S2188
D7S477 D16S3102 HTTINT2 D2S2188 |
MMLS 3.74
MMLS 3.20 MMLS 2.93 MMLS 2.34 MMLS 4.80 |
| Shao | 印刷中 | 96 | 3p
7q Xq |
D3S3680
D7S495 DXS6789 |
MLS 1.51
MLS 1.66 MLS 2.54 |
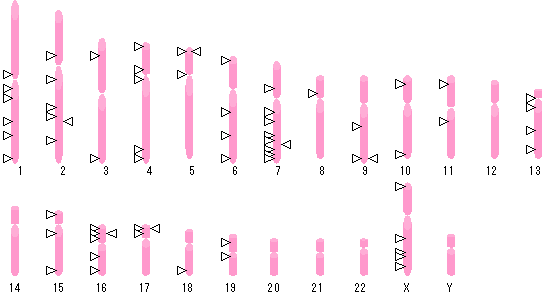
図1 自閉症スペクトルの背景となる遺伝子座のゲノムスクリーン:表1とは異なり,弱いピークも含む.
(オリジナル図を訳者が改変.遺伝子座の位置が一部不正確です)
一連の研究で,X染色体上の遺伝子が自閉症に関与する一つのメカニズムが示された.これは男性により多い他のいくつかの発達障害のメカニズムと同じである.Turner症候群(45, Xカリオタイプ)の対象者を検討し,社会的相互関係の障害は母親由来のX染色体を受け継いでいるケースで非常に高頻度にみられることが,父親由来のX染色体を受け継いでいるケースとの比較で示された.健常の男性は常にX染色体を母親から受け継いでおり,健常女性に比べて,社会的スキルの評価点がより低い傾向が一般的にある.David Skuseは,父親由来のX染色体上で優位に発現する遺伝子座(imprinted locus)がX染色体上にあると想定してこの現象を説明した.ゆえに,母親から受け継いだX染色体は発達早期に表面化する疾患の背景となる常染色体遺伝子座と共に,その(疾患に関連する)常染色体遺伝子座の浸透率を増加させるのかもしれない.
候補遺伝子研究
ゲノムスクリーン研究の公表前に,セロトニン(5-hydroxytryptamine),セロトニン受容体,その他の精神科疾患に関連するとされるトランスミッターなどの代謝に関与する遺伝子から候補が選ばれた.第17染色体上の5-hydroxytryptamine transporter遺伝子はプロモーター(HTTLPR)の中にいろいろな反復配列を持っており,いくつかのサンプルで検討された.最初の報告では自閉症とHTTLPRの短型対立遺伝子の間の関連が報告され,その後,Sabine Klauckらは長型対立遺伝子との関連を報告した.その後3つの研究が長型にも短型にも関連がないと報告し,Nurit YirmiyaらはHTTLPRの長型/長型ホモが長型遺伝子座の優位な親からの伝播と共に自閉症35家系で過剰に見られたと発表した.Sylvie Tordjmanの最近の論文では,発端者における対立遺伝子伝播は障害の重症度に依存しており,重度の障害のケースでは短型対立遺伝子がより多く伝播し,マイルドまたは中等度の障害のケースでは長型がより多く伝播しているとした.したがって,これまでのところ一致する見解は得られていないが,否定的な結果を出した論文は重症度を再検討する必要性が生じている.
報告されている他の異常はdopamine beta-hydroxykase(DBH)の血清レベルが低いことであり,この酵素はドーパミンをノルアドレナリンに変化させる.37組の兄弟ペア家系を,DBH遺伝子のプロモーター領域における19-bpのシークエンスの有無で,DBH+とDBH-にタイプ分けした結果は,有症候兄弟におけるタイプの一致は観察されず,そのかわり,有症候児の母親がDBH-/-遺伝子型を持つ頻度が増加していた.DBH-対立遺伝子は血清DBH酵素活性が低いことに関連していた.著者らは,他のファクターとなる遺伝子型易罹患性とあいまって,何組かの家族における自閉症の背景となっているかもしれない最善ではない(suboptimal)子宮環境の原因に,DBH-/-型に関連する血清DBH酵素活性の低下が関与している可能性を提案している.DBH遺伝子は9q34上にある結節性硬化症遺伝子座(TSC1)に密接にリンクしており,ゲノムスクリーンの結果もこの領域に易罹患性遺伝子が存在することを支持している(図1).
Prader-Willi/Angelman症候群の原因遺伝子座のある15q11-13上の自閉症候補遺伝子は,自閉症ではこの部位での染色体異常の頻度が最も多いために(主に母親由来の重複異常),これまで検討されてきた(表2,図2).自閉症に関連する母親由来の重複異常は通常interstitial duplicationの形を取り,この形の異常はPrader-Willi/Angelman症候群の遺伝子欠損の逆であり,または偽中心体重複(pseudodicentric)性,逆位,重複マーカー染色体ということになる.この15q11-13の重複は,遺伝子量効果を示し,ゲノムインプリンティングの関与を示唆する母親特異性を伴っている.これらのメカニズムのどれかが,あるいは両者が,想定される遺伝性易罹患性の背景となっているのかもしれない.これらの所見から15q11-13上の染色体異常を伴わない症例が検討された.Edwin Cookらは,自閉症とガンマ-aminobutyric acid(GABA)A受容体ベータ3遺伝子の関連を報告した.この所見の再現性は確認されてはいないが,ひとつの研究グループがこの領域に示唆的連鎖シグナルを報告しており,別のもう一つの研究グループが近傍のAngelman症候群の原因遺伝子UBE3A(ubiquitinタンパクligase E3A)との関係を報告している.
表2 第15染色体異常の例
| 著者 | 異常のタイプ | カリオタイプ | 母親からか父親からか |
| Cantuら(1990) | inv dup 15q(女性) | 46,XX,inv dup(15)(q13) | 不明 |
| Kerbeshianら(1990) | del 15q12(女性) | 46,XX,del(15)(q12) | 不明 |
| Gillberg(1991)とMartinsson(1996) | iso 15(男性6例,個別) | 47,XY,iso(15)(pter-q13::q13-pter) | 母親由来 |
| Bundeyら(1994) | interstitial dup 15q11-q13 | 46,XY,dup(15)(q11-q13) | 母親由来 |
| Bakerら(1994) | dup 15q11-13(女性2人) | 46,XX/46,XX,dup(15)(q11.2-q13)
46,XX,+idic(15)(q13) |
不明 |
| Flejterら(1996) | inv dup 15q(個別2例) | 47,XX,+inv dup(15)(pter-q13::p13-pter)
47,XY,+inv dup(15)(pter-q13::q13-pter) |
おそらく母親由来
おそらく母親由来 |
| Hotopfら(1995) | partial 15 tetrasomy(男性例) | 47,XY,+inv dup(15)(pter-q13::q13-11) | 不明 |
| Cookら(1997) | interstitial dup 15q11.2-q12(兄弟2人) | 46,XY,dup(15)(q11.2-q12)
46,XX,dup(15)(q11.2-q12) |
母親由来
母親由来 |
| Chudleyら(1998) | 15qと22qのRobertsonian転座(男性) | 記載なし | 父親由来 |
| Schroerら(1998) | inv dup(15)(男性)
der(15)(男性) interstitial dup 15q11-13(男性) interstitial del 15q11-12(男性) |
47,XY,+dic(15)(q13)
47,XY,+del(15)(q14) 46,XY,dup(15)(q11-q13) 46,XY,deo(15)(q11-q12) |
母親由来
母親由来 母親由来 母親由来 |
| KonstantareasとHomatidis(1999) | inv dup(15)(女性)
der(15)(男性) |
47,XX,+inv dup(15)(pter-q13::13-pter)
47,XY,+der(15)(pter-q15::p11-pter) |
不明
不明 |
| Wolpertら(2000) | idic 15(個別3例) | 47,XX,+idic(15)(q11.2)
47,XX,+idic(15)(q11.2) 47,XY,+idic(15)(q11.2) |
母親由来
母親由来 母親由来 |
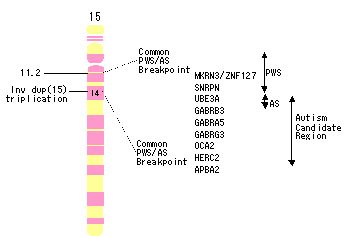
図2 第15染色体領域の自閉症関連候補部位
Prader-Willi症候群(PWS)関連部位およびAngelman症候群(AS)関連部位との関係
APBA2:アミロイドベータ(A4)precursorタンパク結合,family A,member 2
GABRA5/GABRB3/GABRG3:ガンマ-aminobutyric acid(GABA)A受容体,アルファ5/ベータ3/ガンマ3
HERC2:hect domain and RLD2; inv dup, inverted duplication
MKRN3/ZNF127:makorin, ring finger protein, 3
OCA2:oculocutaneous albinism II(pink-eye dilution homologue)
SNRPN:small nuclear ribonucleoprotein polypeptide N
UBE3A:ubiquitin protein ligase E3A(human papilloma virus E6-associated protein)
さらに最近では,候補遺伝子は,連鎖シグナルの位置に近いかどうかと,その遺伝子が脳発達において機能を持つかどうかでリストアップされている.いくつかの研究グループは複数のゲノムスクリーンが指摘した7q22-31の近傍に注目している.また,いくつかの染色体転座症例もまた,候補遺伝子の選択に情報を与えている(図3).検討された候補遺伝子部位はこれまでのところエビデンスの優位性に基づき,自閉症関連遺伝子を含むと最も考えられる部分である120-130cMの部位であった.転座のブレークポイントであるRAY1(現在ST7:suppression of tumorigenicity 7としても知られる)は,自閉症に関する説得力のある(過去の)所見がないままに慎重に検討された.しかし,この遺伝子座に関して新しい所見が報告された.非常に複雑な構造となっており,いくつかの同位体(isoforms)があり,完全に検討されたイントロンの部分にはいくつかのコーディング領域と非コーディング領域が存在する.Thomas Wassinkらは,有症候兄弟例を75組検討し,2例の兄弟ペア家系で有症候兄弟ペアの両者がWNT2(wingless-type MMTV integration site family member 2)に変異を持っていることを発見した.WNT2はRAY1に隣接する.WNT2はいろいろな組織において発達の途中で発現する12個以上のWNT遺伝子の中のひとつで,特に神経系の発達の時期に発現する.さらにWNTシグナルの伝播は,dishevelled(DVL) ファミリータンパクに依存しており,Nardos LijamらはDvl(dishevelled, dsh homologue)のノックアウトマウスにおいて,社会的相互反応の減退を含む表現型を記載した.この表現型は睡眠中に体を丸くする動作の欠如,おりの中の仲間とのグルーミングの消失,母性行動の減少などで特徴づけられる.同じ領域にあるほかのもう一つの遺伝子として,RELN(reelin)があり,この遺伝子は関連研究で自閉症において記載され,Sungmin HongらはRELNにおけるスプライシング変異が常染色体劣性の脳回発達不全の原因になることを同定した.脳回発達不全はニューロンの移動(neuronal migration)の異常のひとつである.

図3 第7染色体の自閉症関連領域
左側は,自閉症発端者で報告された染色体異常.右側はゲノムスクリーンが示唆的連鎖を指摘したマーカーと候補遺伝子の部位.
cM: centiMorgan, del: deletion, dup: duplication, inv: inversion
FOXP2: forkhead box P2
HOXA1: homeo box A1
IMMP2L: inner mitochondrial membrane peptidase 2-like
RAY1/ST7: suppression of tumorigenicity 7
RELN: reelin
SPCH1: speech and language disorder 1
WNT2: wingless-type MMTV integration site family member 2
また,以前SPCH1と呼ばれ,言語発達障害に関連して7q31部位にあることが知られていたFOXP2(forkhead box P2)がクローニングされた.転写因子の遺伝子と考えられるFOXP2内の点変異はKE家系(重症の会話と言語の障害家系:Hurstらが最初に報告)の有症候メンバーにおいて発見された.KE家系と血がつながっていない一人の(言語障害)患者の転座ブレークポイントは,このFOXP2にあることも指摘された.自閉症と重症の言語障害は7qのこの領域にある関連遺伝子を共有している可能性もある.まとめると,7q領域は自閉症易罹患性遺伝子を探している研究者たちにとっては,引き続き最重要部位ということになる.
候補遺伝子研究の最後の例としては,Stephane Jamainらが6q21にあるグルタミン酸受容体ionotropic kainate 2(GRIK2:GLUR6としても知られる)遺伝子の多型と自閉症との間の連鎖の有意な証拠を発見している.この遺伝子は位置的にも機能的にも候補遺伝子として有望である.33例の有症候者における変異スクリーニングでは,いくつかの単一塩基多型が明らかにされ,GLUR6での単一アミノ酸変化を伴う変異を含んでいた.この変異部は高度に変異の少ない部分であり,自閉症者の8%にこの変異が見られた.この多型の機能的意義はまだ明らかになっていない.
他の原因
多くの自閉症ケースは,提唱されている複数遺伝子性とエピスターシス(遺伝子間の相互作用)のメカニズムに矛盾しないが,少数例では染色体異常や結節性硬化症などの特別な原因を持つ疾患に関連している.環境因子もまた考慮されているが,説得力のある証拠はほとんどない.
染色体異常: 自閉症スペクトル者における染色体異常の報告は,ほとんど全ての染色体に関してなされている.常染色体の倍数体でない染色体異常(aneuploidies),性染色体異常,欠損,重複,転座,リング状染色体,逆位,そしてマーカー染色体異常などがある.自閉症における染色体異常の予想頻度は5%から48%と報告により巾があり,基本的には対象者のIQが非常に低いかあるいは身体奇形ケースが含まれているかどうかに依存している.特発性自閉症に関する研究では,染色体異常の頻度は通常5%以下である.最もよく見られる染色体異常は前述したように第15染色体のPracer-Willi/Angelman症候群領域の異常である.
既知の医学的状態: 一般的に自閉症者10%から15%は,メンデルの法則に従う検出可能な遺伝性の症候群(染色体異常を含む)を持っていると言われている.文献で報告されているそれらの多くは症例報告であるが,いくつかの状態はいくつかの異なる研究グループによって体系的に検討されている.
自閉症とメンデルの法則に従う遺伝性疾患の関連の報告で,最も古いのはフェニルケトン尿症に関するものである.Folsteinは,自閉症とフェニルケトン尿症の関係についてのエビデンスをレビューし,初期の頃のレポートは信頼できる自閉症の診断基準に基づいているのかがはっきりしていないため,過去の研究の解釈は困難であると強調している.おもしろいことに,幼児以後実際に食事療法が開始されると自閉症症候は多くの場合消失するが,IQの改善は見られない.1980年代に自閉症スペクルである対象者にフェニルケトン尿症患者がいないかどうかのスクリーニングが行われたが,一致した結果は得られていない.しかも,各サンプルサイズが小さいため結果の解釈は困難である.幸いなことに,未治療のフェニルケトン尿症はイギリスでは新生児スクリーニングが普及し無視できるほどの数になり,そのためにこの件に関する研究は不可能となっている.
脆弱X症候群,結節性硬化症,そして神経線維腫症は,自閉症と関連があるメンデルの法則に従う状態として文献では頻回に記載されている.自閉症者における脆弱X症候群の有病率は,論文により大きく異なっており,0%から20%と報告されている.同様に脆弱X症候群患者の中の自閉症の頻度は5%から60%と報告されている.このような報告間の差はおそらくサンプルサイズが小さいことや,評価方法の違い,そして診断や脆弱X症候群の検査方法の不一致などによると考えられる.さらに,多くの論文が脆弱X症候群のDNAテストが可能になる前に行われており,脆弱X染色体の診断は染色体検査で行っており,信頼性も低く,特異度も低い.
自閉症者の中での結節性硬化症の頻度は2つの疫学的研究において0.4%から2.9%と報告され,3番目に行われた研究では14%と有意に多い結果であった.この3番目の研究ではけいれん発作を伴っているサブグループが含まれていた.結節性硬化症における,自閉症や,典型的でない自閉症や,広汎性発達障害の有病率もまた論文によって差があり,20%から61%と報告されている.結節性硬化症における自閉症発生率は脳における結節の位置に関連している可能性が示唆されている.自閉症者の中での神経線維腫症の推定頻度については3つの報告があり,その結果には0.2%から14%の巾がある.この差はおそらく,身体所見,MRI所見,または過去にさかのぼったカルテのレビューなどにおける検討方法の違いによるものであろう.
最近,Duke大学の研究者たちは,Rett症候群ではない自閉症者において,MECP2(methyl CpG-binding protein 2:Rett症候群の変異遺伝子)に変異を発見した(未発表).Rett症候群ではなく,自閉症と診断された69例の女児例から,2人がMECP2遺伝子に変異を持っていたのである.このことは,Rett症候群が以前考えられていたよりもさらに非単一性を持っていることの証拠なのかもしれない.
環境因子: 自閉症の発生率が明らかに増加しているのは,主に環境因子によるとする理解が,一般大衆に広がっている.しかし,発生率の増加の大部分は診断基準の緩和と,この基準を正常知能の人々から非常に知能の低い人々や,医学的(他の)異常や染色体異常を有するケースにまで広く適用したためであるようである.にもかかわらず,メディアは環境原因の理論にかなり注目しており,そのようなメディアにとっては支持的なエビデンスがいくつか存在する.
最初の頃の論文の多くは,主に症例報告で,自閉症と母親の甲状腺機能低下症の関連,自閉症と先天性甲状腺機能低下症の関連,自閉症と母親のサリドマイド服用との関連,自閉症と母親のバルプロ酸使用の関連,自閉症と母親の飲酒の関連,そして自閉症と先天性サイトメガロ感染症の関連などが記載された.最も説得力のある報告は,先天性風疹症候群と自閉症の間の因果関係に関するエビデンスで,243例の症例検討を基盤にしている(表3).このうち22例が典型的あるいは非典型的自閉症を呈している.Murdina Desmondらは,64例の先天性風疹症例の中で8例が自閉症であったと報告した.
表3 自閉症において提唱された環境因子
| 可能性のある因子 | 報告者 | 方法 |
| 母親の甲状腺機能低下症 | GillbergとKopp(1992)
Haddowら(1999) |
症例報告:2例
thyrotropinレベルが上昇していた母親62例の子供の認知検査 |
| 先天的甲状腺機能低下症 | GillbergとKopp(1992)
RovetとEhrlich(2000) |
症例報告:3例
先天性甲状腺機能低下83例の認知検査 |
| 母親のサリドマイド内服 | MillerとStromland(1993)
Stromlandら(1994) |
サリドマイド胎児期異常とされていた86成人例を追跡研究
上記の追加検討 |
| 母親のバルプロ酸使用 | Christiansonら(1994)
WilliamsとHersh(1997) |
症例報告:2例(兄弟)
症例報告:1例 |
| 母親の飲酒 | Nanson(1992)
Aronsonら(1997) |
症例報告:6例
妊娠中飲酒ケースの26例の追跡研究 |
| 先天性サイトメガロウイルス感染症 | Stubbs(1978)
Stubbsら(1984) |
症例報告:1例
症例報告:2例 |
| 先天性風疹感染症 | Desmondら(1970)
Chess(1971) Chess(1977) |
先天性風疹症候群64例の発達評価
先天性風疹症候群の243例の行動研究 上記のフォローアップ |
| MMRワクチン | Wakefieldら(1998)
Taylorら(1999) Patjaら(2000) Kayeら(2001) |
明らかな広汎性発達障害と消化管症候の合併(12例)
イギリスにおける498例の自閉症ケースの疫学研究 MMR接種後に重症な副反応を起こした例の追跡研究(フィンランド) 時系列解析(イギリスのデータベース) |
自閉症の原因として広く一般大衆に注目された最新の環境因子は,MMRワクチンである.この大変深刻な懸念に対して短期間で集められたデータの大半は因果関係を支持していないが,この件をとりまく世論は自分の子供にワクチン接種を受けさせない風潮を招来している.この関連の可能性は最初にAnthony Wakefieldらが提唱した.彼らは明らかな発達退行と消化管症候を伴う12人の小児を報告した.12例中8例で,親は行動症候のオンセットとMMRワクチン接種が関連していると述べた.伝えられるところによれば,12例中9例は自閉症的で,いったん発達した言語とスキルが明らかに失われた症例である.しかし,MMRワクチン接種前の機能レベルに関しては詳細が伝えられておらず,脆弱X症候群のスクリーニングは受けているものの,形態異常の検査や染色体異常の検査はいずれも記載されていない.著者ら自身は,MMRワクチン接種と自閉症の間の関係を証明したわけではないとしており,1988年にMMRがイギリスに導入された後,自閉症の発生率が増加しているのかどうかに結論を得ることが重要であると主張している.
Brent Taylorらは,1979年から1992年の間にイギリスで生まれた自閉症児に関して,自閉症の頻度とMMR導入時期との関係を検討する研究を行った.スクリーニングにより498人の自閉症児が同定され,そのうち293例は,典型的自閉症か非典型的自閉症あるいはアスペルガー症候群であることが確認された.出生年齢別では,自閉症ケースの増加は一定で,1988のMMRの導入の後の急変はなかった.18ヶ月前後にワクチンを受けたケースとワクチン接種を受けていないケースの間では,診断年齢に差はなく,自閉症のオンセット(MMR接種時期の1-2年後)との間に時間的関連は見いだせず,また発達退行現象も接種後の数ヶ月に集まっているわけでもなかった.
双生児研究で一卵性一致率が100%でないとすると,環境因子が自閉症の発生率に寄与している可能性はある.その場合はおそらく遺伝的な易罹患性を持つケースにおける「セカンド・ヒット」としての作用であろう.しかし,一部の少数のケースでは環境因子が主な原因となる可能性もあるが,大多数のケースの主な原因と成り得るような単一の環境因子は報告されていない.あるいはまた,一卵性双生児における表現型の双生児間の多様性もまた,確率的な因子(環境因子)の結果なのかもしれない.
遺伝カウンセリングと教育
自閉症に関する遺伝カウンセリングは,表現型の複雑性と背景となる原因の複雑性のために大変難しい.Emily Sinonoffは家族歴の慎重な評価と発端者の評価が必要であることを強調している.兄弟内再発率や将来の妊娠時の出生前診断の可能性に関する正確な情報を提供するためには,メンデルの法則に従う状態や症候群や染色体異常を除外することが肝心である.理想的には,自閉症スペクトルと診断を受けたケースの評価には,医学遺伝学者または身体形態異常に関して臨床経験をつんだ臨床医の診察が含まれるべきである.しかし,身体形態異常が明らかにない場合でも,分子レベルの検査や染色体検査が必要でないわけでなない.例えば,脆弱X症候群では身体形態異常が全くない例も存在することがよく知られている.従って,自閉症スペクトル者における遺伝学的検査は,少なくとも,高感度染色体検査と脆弱X症候群の検査を含んでいるべきである.熟練した臨床医によるWoodランプテストも結節性硬化症のような神経皮膚症候群の可能性を評価するために必要である.
特発性自閉症の場合,遺伝カウンセリングは自閉症の遺伝の複雑性についての議論を含まなければならない.(特発性自閉症の)遺伝学的検査はまだ不可能で,もし複数遺伝子遺伝というモデルが正しいとすると,易罹患性遺伝子が同定されたとしても特発性自閉症の遺伝学的検査はできないかもしれない.検査できるとしても,現在可能になったアルツハイマー病の検査に類似したものになるであろう.ゆえに,出生前診断にはほとんど役に立たないであろう.特発性自閉症の兄弟内再発リスクを評価する際には,臨床家は経験的なリスク値から判断せねばならない.兄弟における有病率はどの報告でも一致して3%までであるが,この値は「産み止めルール」のために過小評価した値である.発端児が小さいころから重症のハンディキャップを有している状態では,親の出産計画に影響し,次の子供をつくらないか,あるいはつくっても一人である.自閉症発端者に続いて生まれた兄弟のみを検討した場合,自閉症の兄弟内発生率は6%から8%になってしまう.家系内に2人の自閉症児がいる場合,三番目が自閉症であるリスクは25%に及ぶとする未発表の見解もある.より広汎な自閉症表現型のひとつ以上を有するかどうかのリスクは成人兄弟内で30%と報告されている.
自閉症の遺伝カウンセリングに関する文献はなく,実際行われているカウンセリングの方法はさまざまである.家族は自閉症の遺伝学的情報についてはあまり説明を受けていないが,知りたいという意欲は旺盛である.基本的遺伝原則や特に自閉症の遺伝学に関する専門的教育もまた必要とされている.我々はDuke大学の共同研究者と合同で,インターネットを基盤とする家族のための教育ツールを開発し始めた(http://www.exploringautism.com).遺伝学の専門家と自閉症の臨床家の両方のための教育プログラムにつながる研究努力も開始している.
展望
自閉症の原因となる遺伝子がもし発見されたら,自閉症を予防したり治療したり重症度を軽減することにつながるかもしれない介入方法が発見されることを期待して,その発現のタイミングや作用メカニズムが研究できるようになるであろう.ここでは,いくつかの相補的なアプローチを記載する.そのいくつかは,大きなサンプル数を得るために,研究者達がそれぞれのサンプルを提供して集めることを必要とするアプローチである.
臨床的多様性の利用: この戦略は二つのステップを含んでいる.一つ目は遺伝学的研究のためにはサンプルを発端者がKannerが記載したような自閉症者の家系に制限することである.Kannerはたとえ行動学的基準は満たされていても,身体形態異常や既知の原因を持つケースを除外した.
これらの「クラシカルな自閉症家系」における表現型の多様性の十分な利用に必要な二番目のステップは,量的な側面でも質的な側面でも全ての側面で発端者の特徴を慎重に検討し,同様に両親と自閉症でない兄弟に関しても自閉症発端者の家族メンバーに多いことが判っている症候に関して慎重に検討することである.しゃべることがない重症の発端者から知的レベルが正常範囲に入っている発端者まで多様である.例えば,社会的な行動や衝動的な行動の重症度においても多様である.また,多動,ある種の音や触覚や場合によっては味への過敏,神経心理学的テストの結果,そして顔の表情などのような診断基準に含まれていない多くの所見に関しても多様である.
両親の特徴としては,言語や語用(プラグマティクス)あるいは管理統合能力(executive function)における異常,IQパターンの異常,社会的寡黙性,そしてルーチンへの融通の効かない執着と変化を嫌う傾向などが含まれる.その他には,頭が大きいこと(macrocephaly)や短頭(brachycephaly:頭蓋骨の前後径が短く,その結果比較的横径が大きくなる),血中セロトニンの増加,気分障害(mood and anxiety disorders)などがある.これらの形質は,確かに異常と思われるケースに伴うこともあるが,一般ポピュレーションにも存在しており,必ずしも機能的障害に関連しているわけではない.しかし,自閉症の最も有望視されている遺伝モデルである遺伝子間の相互作用を伴った複数遺伝子性モデルからすると,寄与している遺伝子は一般ポピュレーションにおいても一般的なものである必要があると想定される.家系のケース・コントロール研究で浮き彫りにされる症候の中で,自閉症家系メンバーの3分の1以上で見られる症候は皆無である.従って,特定の形質に関する遺伝解析を行うために十分な家系数を得るためには,大きなサンプルサイズが必要である.
この理由から,いくつかの研究グループは巨大なサンプルを集めるための試みの中で共同研究を始めている.最も大きな共同研究は,自閉症遺伝学共同体(Autism Genetics Cooperative: AGC)で,以前は別々であった8つのグループからなり,集められたサンプルは800組の兄弟ペア家系である.AGCは参加するだれもが入手することができる共通データベースの中に全ての臨床データやマーカーデータを集めるという大変な企画を遂行するために研究費を申請した.これまでのところ,北アメリカでの兄弟ペア家系収集のための家族を基盤とする団体である,自閉症遺伝リソース交換(Autism Genetics Resource Exchange: AGRE)は,197組の複数発生家系からの自由に入手可能なDNA情報および信D何情報を保有している.もう一つの共同研究グループは200家系以上のデータを持つ,国際分子遺伝研究自閉症会議(IMGSAC)で,いくつかのイギリス,ヨーロッパおよび北アメリカの研究者たちが共同研究を行っている.
異なる構造を有する家系: これまでに行われた連鎖研究は,2人あるいはそれ以上の有症候児を持つ家系をサンプルとして使っていた.Neil RischとZhangによって提唱されたもう一つのアプローチは,極端に不一致の兄弟ペアをサンプルとして使う方法で,この方法では,自閉症発端者とその兄弟で完全に自閉症傾向を欠く者を比較する.そのような家系はあまり一般的ではないが,この方法は理論的には小さい効果の遺伝子を検出するためには強力な方法である.また,いとこ関係で,両者が自閉症である場合は有症候兄弟ペアの場合よりも共有する遺伝子の数が少ないためバックグランドノイズが少なくなり,自閉症関連遺伝子検出には強力であろう.
このアプローチを拡大したものが,巨大一族の研究である.このような一族はほとんど知られていないが,我々が知っている範囲では,下記の例を除き,十分な解析に耐え得る十分な数の一族を集めることに成功した研究者はいない.このただ一つの例外は,ユタ州に在住する一族で,ユタ州のユニークな家系図情報を使い,いくつかの核家族例の祖先をさかのぼって検討し,共通する祖先をつきとめたものである(未発表).
組織サンプル: ヒトの脳組織が研究できれば,神経学的障害における研究には非常に有用である.このアプローチはその障害に関連する神経化学システムを同定することにより候補遺伝子のヒントを得ることもでき,一旦遺伝子が見つかった場合にはメカニズムを研究するための決定的な研究素材を提供する.神経科学的アプローチは,自閉症者およびコントロール者の脳組織が少ないために遅々として進まない.自閉症者の寿命は正常であるので,死体解剖プログラムを体系化することは困難である.10年前までは,脳組織の献体を求めていたのはMargaret Baumanだけであった.1992年からは,イギリスで献体募集が実施され,4年前には自閉症研究のための全国同盟(National Alliance for Autism Research)が北アメリカでの先鋒となり,アメリカ自閉症協会(Autism Society of America)とDavisにあるCalifornia大学のM.I.N.D.研究所と共同で自閉症組織プログラムを結成した.これまでのところ,自閉症組織プログラムは28例の献体を得(自閉症者およびコントロール),18人の研究者に組織検体を供給している.
動物モデル: 自閉症に関連する変異やバリアントを持つ遺伝子が発見された場合,もちろんそれらの遺伝子はげっ歯類でも検討することができる.しかし,ヒト以外の種を研究している科学者にとって同様に重要なのは,社会性や反復行動や特定の認知症候における多様性に関心を持つことであり,ヒト以外の種の行動の多様性を自閉症の遺伝学や神経科学を研究している科学者たちに知らしめることである.これまでのところそのような研究で最も重要なのは,Thomas Inselらのプレーリー地区のハタネズミと山地のハタネズミの間の社会性の違いに関する研究である.この二つの種は,片方が社会的で一夫一婦主義であるのに対し,もう片方は単独行動性で無差別結婚主義であり,バゾプレッシンとオキシトシン遺伝子のプロモーター部に違いがある.この遺伝子の違いがニューロン移動のタイミングに影響し,その結果脳の中でのこれらの遺伝子の局所的発現が異なってくる.
結語
この10年足らずの研究を基盤にして,遺伝素因はあきらかに最も有力な自閉症スペクトルの原因として取り上げられるようになった.染色体異常やメンデルの法則に従う特異な疾患との関連に関する症例報告は数多くあるけれども,おおくのケースは特発性で明らかに複雑な遺伝パターンを背景としている.このことが易罹患性遺伝子の発見を困難にしている.にもかかわらず,いくつかの遺伝子(座)に関して注目される染色体領域を同定することに関しては,かなりの進歩があり,特に第2染色体,第7染色体,第15染色体,X染色体に関しては報告が成された.これまでのところ所見は決定的ではないが,候補遺伝子に関する研究は進行中である.候補遺伝子の選別は,連鎖シグナルの近傍にあることや,染色体異常のブレークポイントにあること,胎児発達時期の脳における発現性,そして限られてはいるが病態生理学的知識に基づいている.補足的アプローチには,動物モデルやヒト脳組織の研究がある.自閉症の遺伝に関するより意義のあるデータを得るためには共同研究が必要であるとする認識に加え,このような強力ないろいろなアプローチ法がそろっているので,今後の革新的な進歩が期待でき,この最も難解な状態(自閉症)を理解できる日が近いという楽観的な考えを持つことができる.