まとめ:会話と言語の特異的発達障害(SDDSL)は,表出および(または)受容性の言語能力の発達障害で,知的障害,感覚異常,肉体的障害,神経学的障害によるものではない.家族研究と双生児研究の結果は,自閉症においてもSDDSLにおいても,遺伝素因が重要であることが示されている.自閉症者の親族にはSDDSLが多いことが知られており,SDDSLが自閉症の遺伝素因が関係した一つの発現型である可能性も示唆されている.SDDSLを自閉症の軽症発現型と考え,自閉症とSDDSLの発症率をモデル化すると,3つか4つの遺伝子座が相互作用を持つ形で(epistatic loci)関与していることが示唆されている.最近発表された連鎖研究では,明らかに優性遺伝のSDDSLの家系が報告され,7q31が易罹患性遺伝子座の候補としてSPCH1と名づけられた.この部位はまた,自閉症の複数発症家系においても連鎖が報告されている.我々は,この論文で,7q31.3部のブレークポイント(切断・結合点)を含む遺伝子再配列を呈する自閉症例とSDDSL例を提示する(異なるタイプの再配列).また,FISH法(fluorescence in situ hybridisation)を使い,ブレークポイントの場所を確認した(CFTRとD7S643の間の1cM以内の部分).我々の結果は,自閉症の遺伝素因にも関連し,SDDSLや自閉症の易罹患性遺伝子座の今後の研究にとって重要なものである.
(イントロ)会話と言語の特異的発達障害(SDDSL)は,特異的言語障害(SLI)を含む概念である.SDDSLと自閉症および他の広汎性発達障害と診断される児において,形質上の重複の有無に関してはすでに長く議論されている.SDDSLにおいても自閉症においても,家族研究と双生児研究は遺伝素因の存在を示唆している.
最も最近のイギリスでの自閉症に関する双生児研究では,自閉症の遺伝性を91-93%と想定している.自閉症の双生児不一致例の中で,特異的言語障害を含む特異的認知障害が,一卵性の不一致例の自閉症でない方の児の50%,二卵性の不一致例の自閉症でない方の児の10%に報告されている.これらのデータは,家族研究の結果と共に,自閉症の遺伝素因の発現型の一つとしてSDDSLがあり得る可能性を示唆している.
SDDSLの大規模双生児研究は,SDDSLの二つのサブタイプ(表出性言語障害,発音障害を伴った受容性言語障害)の遺伝性がかなり高いことを示し,SDDSLとより特異的でない障害(非言語性能力低下例あるいはボーダーライン例における言語障害)の両者の遺伝的背景が同じである可能性も示唆した.より全般的な学習障害の例がSDDSL児の中に存在しており,また自閉症児の80%が知的障害を有している.
自閉症やSDDSLの背景となる神経病理学的基盤や,両者の関係に関しての結論はでていない.家系内に最も多発している家系でも遺伝形式は不明瞭である.SDDSLの多発家系における遺伝形式も通常はメンデル型の遺伝パターンではない.しかし,SDDSLではKE家系という巨大家系が知られており,常染色体優性遺伝形式である.有症候者は,言語および口・顔面の統合運動障害と言語障害を呈し,受容性言語よりも表出性言語の方がより障害されている.広範囲な認知障害も存在し,結果的にIQは下がって境界部に位置する.有症候者ではまた,脳のいつくつかの部位で機能的・構造的異常がみられると報告されている.
KE家系における連鎖研究では,第7染色体の7q31に強い連鎖が見つかっている.複数ポイントマッピングでは,想定される関連遺伝子SPCH1(ロッドスコア3)は,マーカーD7S2459とD7S643の間で,5.6cMの区間に存在することが示唆された.マーカーD7S2425とCFTR(嚢胞性線維症膜透過コンダクタンスレギュレーター)遺伝子の間の3.8cMの区間が複数ポイントロッドスコアの最高値(6.62)を示した.近隣のいくつかの遺伝子の微小欠損の可能性を否定することはできないが,報告者らは透過率の高い(full penetrance)単一の遺伝子座がSDDSLの原因遺伝子であることを推測した.SDDSLと7q31のマーカーの関連についての研究の予報では,SDDSL児の拡大サンプルにおいてはSFTR遺伝子とSDDSLとの間に関連が示唆されている.
最初のゲノム全長に渡る自閉症連鎖研究の結果(自閉症分子遺伝研究会議:IMGSAC)は,自閉症の関連遺伝子は第7染色体上にあることを示唆した.有意な連鎖を示した遺伝子部位はD7S527とD7S483の間(7q22-7q35)で40cMという大きな区域であるが,7q31-35におけるマーカーD7S530とD7S684の近傍に複数ポイントロッドスコア2.53というピークが見つかった.このIMGSACの論文では,自閉症形質の定義を厳密に設定し,DSM-IVやADI-Rなどの基準に必要な状態よりも言語障害がよりはっきりしている症例を選んで検討している.
自閉症における他の連鎖研究でも,7q3に関して陽性所見を報告している.Philippeらは7q31部位のD7S486に複数ポイントロッドスコアのピーク(0.83)を報告し,Ashley-KochらはD7S2527に複数ポイントロッドスコアのピーク(1.77)を報告した.RischらもD7S1804にピーク(0.93)を報告している.
自閉症に関する全ての連鎖研究がまた,他の遺伝子領域に関しても陽性所見を得ており,自閉症関連遺伝子の候補部位が他にもあることが示唆されている.さらに,自閉症の中に脆弱X症候群や結節性硬化症などと関連する症例があり,自閉症の病因的多様性が予想されている.
臨床像および染色体所見
症例1:出生時異常なく,親の血族結婚もない.初期発達は遅れがあり,17ヶ月で歩行開始.最初の臨床評価は,20ヶ月時で,Griffiths発達スケールで11-12ヶ月に匹敵し,McArthurテストでの言語とジェスチャーは8ヶ月レベル以下であった.ICD-10では,自閉症の診断基準を満たした.31ヶ月時の最近の評価では,非言語性IQは22ヶ月に相当し,発語はなく,ADI-RでもADOSでも自閉症診断基準を満たした.従って,臨床プロフィールは,発達の全ての面で明らかな遅れがみられ,その中でもコミュニケーション能力と行動に関しては特に遅れており,自閉症の診断基準を満たす状態であった.母親の兄弟(本人のおじ)は,アスペルガー症候群で,いとこにSDDSLが一人存在する.染色体検査では,第7染色体動原体近傍の逆位がみられた.
症例2:妊娠中の異常なく,38週で正常分娩.親に血族結婚なし.新生児期の異常なし.運動機能の発達正常.7ヶ月で座り,11ヶ月で歩いた.2歳までに数単語しかしゃべらず,2単語の連結は3.5歳で可能となった.4歳5ヶ月時の耳鼻科的診察で聴力に問題なし.よちよち歩きの頃から活発で,反抗的行動みられ,保育園では同年齢の友達との関係が困難で,通常の小学校では,常時補助の先生を必要とした.5歳の時反抗挑戦性障害と診断され,はずかしがりやであることと情緒的未熟さをも指摘されている.4歳10ヶ月時のGriffiths発達スケールでは,正常範囲の下限.5歳時のReynell発達言語スケールでは,文法は正常であったが,表出言語および会話発声に障害がみられた.5歳5ヶ月時のWeschler就学前知能スケール(WPPSI)では,言語スコアが低く10ポイントの解離がみられた(有意差なし).全般IQは74.6歳3ヶ月の時の基礎言語臨床評価(CELF-R)では受容性においても表出性においても5歳以下と判定された.6歳8ヶ月時でも,学校での言葉による指示には従えず,ボキャブラリーは十分あったが,考えを表出するのは困難.単語を見つけたり,配列したり,明瞭に発音することが困難.ADI-RとADOSでは広汎性発達障害の基準は満たさず,利き腕は右,脳波正常,6歳11ヶ月時のMRI検査では右大脳半球の側頭葉角部に小さな異常(dysembryoplastic neuroepithelial tumour疑い)を指摘された.肉体的な発達は正常で,全般IQは正常下限であることから,言語発達の障害とされた.また,注意力と行動上の問題も指摘された.母親は躁うつ病の既往があり,母方に言語発達の遅かった人がいたらしいが,未確認.第2染色体と第7染色体の間に相互転座異常がみられた.
方法
染色体検査:末梢血リンパ球をPHAで刺激培養後,GTGバンド法でメタファーゼ期の細胞を解析した.
分子レベルの染色体検査:7q31.3におけるブレークポイントは,FISH法(fluorescence in situ hybridisation)にて場所を同定した.使用したプローブは,YACクローン法で作成し,KE家系研究で最も連鎖の高かった遺伝子マーカーを含んでいる.YACクローンはD7S643(881-c-6)とD7S633およびD7S677(751-e-9)を含んでおり,D7S677とCFTR遺伝子(エクソン24)を含むクローンHSC7E127(プローブ)は,S. Scherer先生とS. Tosi先生からいただいた.FISHプローブはDaviesらの方法で処理し,市販されているBio-NickキットでNick translation(断片化したラベリング)を行った.Daviesらのプロトコールは,市販でないプローブのhybridizationと検出に使用.市販プローブ(D7S486)は,説明書に従って処理した.
結果
D7S486と751-e-9(D7S633とD7S677)とHSC7E127(D7S677とCFTR遺伝子エクソン24)と881-c-6(D7S643)でFISH法の結果が得られた.これら全てのマーカーは,第7染色体の先端から累積距離で145cMの位置にある.HSC7E127と881-c-6の二つは,両者の間(1cM以内)に7q31.3のブレークポイント(切断点)があることを示した(2例とも).このブレークポイントの位置は,連鎖研究の結果に矛盾しない.KE家系での結果でも,複数ポイントロッドスコアのピーク(6.62)は,D7S2459(140cMの位置)とD7S643(145cMの位置)の間である.145cM部位のマーカー(D7S486とCFTR)は,それぞれ最大2点ロッドスコアが6.22と5.46と報告されている.Phillippeらの自閉症に関する検討では,D7S486(145cMの位置)で複数ポイントロッドスコアのピーク(0.83)を得ている.Ashley-Kochらは,自閉症に関する検討で,151cMの位置(6cM以内遠位側)のD7S2527で複数ポイントロッドスコアのピーク(1.77)を報告し,IMGSACは,D7S530とD7S684の近くで,上記のブレークポイントの12-24cM遠位側に連鎖のピークを報告している.また,Rischらの検討でも,IMGSACが報告したピークの10.3cM近位部のD7S1804にピークをみとめている.
考察
症候に類似性がある二人の患者において,両者とも7q31.3部位での1cM以下の範囲内にブレークポイント(切断・結合点)のある遺伝子異常がみられたことは興味あることである.ブレークポイント部で遺伝子機能を破壊する遺伝子再配列は,疾患遺伝子の同定に多大に寄与してきた.7q31.3部のブレークポイント部の役割が不明であっても,他の支持的な遺伝データの存在により,関連が疑陽性となる可能性は少なくなる.
ここで報告した2例のブレークポイント7q31.3は,これまでに発表された連鎖研究で指摘された関連遺伝子の位置と驚くほど一致している.3つの研究報告が,このブレークポイントから6cM以内に連鎖のピークを報告している.また,IMGSACの報告では,CRTRとD7S486では陽性結果がでて,連鎖のピークはこのブレークポイントから12-24cM遠位側にあった.これまでの他の例では,関連遺伝子の部位とブレークポイントの位置には複雑な関係が存在することもあり,関連遺伝子から1Mb離れたところに再配列のブレークポイントがあった例もある.従って,IMGSACが報告した連鎖ピークとブレークポイント位置とのずれは,両者が7q3にある単一の関連遺伝子の存在を示唆している可能性を否定するものではない.あるいは,7q31.3より遠位側にある遺伝子座が連鎖結果に影響しており,7q31.3のブレークポイントはもう一つの関連遺伝子の位置を示している可能性もある.また,単なる疑陽性結果の可能性も残る.
自閉症とSDDSLは臨床的には関連が強く,1cM以内にある2症例のブレークポイント位置が,KE家系のSDDSL関連遺伝子(SPCH1)に近接していることは興味深い結果である.自閉症症例(症例1)とのこの位置関係は,共通の遺伝子座(SPCH1である可能性あり)が,SDDSLと自閉症の両者の病態に関係している可能性を示唆する.もちろん,密接に関連するか近接した遺伝子が,異なる形質にそれぞれ関与している可能性は否定することはできない.
最近報告された母親から伝わった第7染色体逆位の家系で,自閉症とSDDSLの両者に7q31.2(7q31.3の近位側)が関わっていることが示唆された.この家系の二人の兄弟(男性)は自閉症であり,妹は著明な発達遅滞と表出性言語障害を呈していた.この家系では,遺伝の複雑さを説明するためにインプリンティング(発現が母親由来か父親由来かで異なる)が想定された.また,このブレークポイント(7q31.2)が付加的な意味を持つことは否定できない.
我々の結果と,これまでに報告された結果は,7q31に自閉症とSDDSLの関連遺伝子がある可能性を示唆し,第7染色体上のこの部位の中か近傍に,一つあるいは複数の関連遺伝子が存在する可能性を示唆する.
症例1の家系内発現パターン(自閉症,アスペルガー症候群,SDDSL)は,自閉症の遺伝研究結果に矛盾せず,いくつかの遺伝子座がお互いに影響し合っていることを示唆する.自閉症複数発症家系における再発生率の数学的モデリングは,複数遺伝子座モデルを示唆しており,Picklesらによると3つの遺伝子座が最適で,10個まであり得るとしている.Rischらは,さらに多い関連遺伝子の可能性を指摘している.症例1の母方の家系に軽症型ともとれるアスペルガー症候群とSDDSLの例が存在しており,逆位のない第7染色体の同じ部位での変異を含む一つあるいは複数の他の関連遺伝子が母方家系に伝わっており,それに7q31.3部位での変化(逆位によるブレークポイント)が加わって自閉症者が発生していると考えることもできる.母方のおじがアスペルガー症候群である点は,社会的・行動的側面と言語コミュニケーションの側面が自閉症においては,別々の関連遺伝子で支配されている可能性をも示す.
症例2の右大脳半球の腫瘍については,左半球ではないので言語障害に関してはその原因とは考え難い.
自閉症発端者の親族に,うつ病や社会恐怖症が増えているという報告がある(1999年のPivenとPalmerの論文は否定).症例2の母親は躁うつ病の既往があり,一つあるいは複数の関連遺伝子が存在している可能性が示唆される.しかし,発現形質の定義は難しく,SDDSLは年齢によって様相が異なり,成人における言語障害の評価は困難であるため,少なくともいくつかのSDDSLに影響する単一のメジャーな関連遺伝子の存在を否定することはできない.故に,症例2のSDDSLは,単に優性に作用する7q31.3ブレークポイントの結果であるのかもしれない.
Bishopらは,SDDSLの双生児研究を行い,遺伝性の高いSDDSLのサブタイプとして「AEサブタイプ」を提唱した.AEサブタイプは,発音障害と表出言語障害で,受容性言語障害はあってもなくてもよいサブタイプとされた.症例2とKE家系症例の両方の形質をこのAEサブタイプと考えることも可能である.
この報告は,1cMの範囲内で7q31.3部位にブレークポイントを有する二つの遺伝子異常を報告し,自閉症とSDDSLの病態における連鎖研究の結果と比較して検討した.このブレークポイントは,特に,自閉症とSDDSLの両者の病態に共通の遺伝子座(おそらくSPCH1)に関連しており,これらの疾患の関連遺伝子を探す研究において重要な候補となる.
(解説)先日もAshley-Kochらの論文(文献1)の解説で,第7染色体についてまとめましたが,この論文とBarrettらの論文(論文2)を追加して,さらに細部に関してまとめてみます.
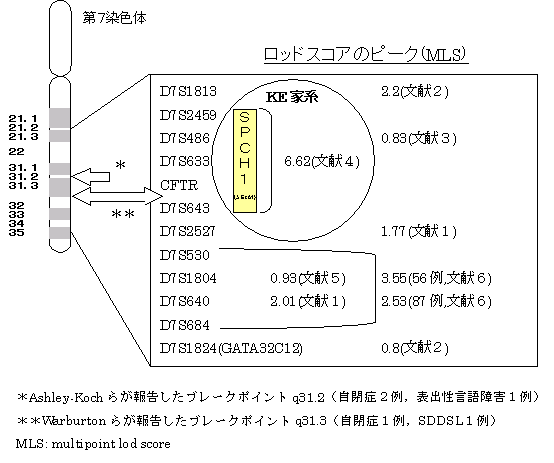
会話と言語の特異的発達障害(SDDSL)の優性遺伝家系と言われるKE家系の関連遺伝子候補部位SPCH1の周辺に,自閉症研究でのロッドスコアのピークが散在しています.